το ΘΕΜΑΤΟΛΟΓΙΟ ΠΝΕΥΜΟΝΟΛΟΓΙΑΣ ουδεμία ευθύνη φέρει για ενδεχόμενες διαγνωστικές ή θεραπευτικές αστοχίες των αναγνωστών του
♦♦►ΕΠΙΚΑΙΡΑ ΑΡΘΡΑ
ICD-10
 Πόλεμος, Κρίση, κλιματική αλλαγή, μετανάστευση και αναπνευστική λειτουργία
Πόλεμος, Κρίση, κλιματική αλλαγή, μετανάστευση και αναπνευστική λειτουργία
επιλεγμένα θέματα
H Tεχνητή Νοημούνη στην Ιατρική
Καρδιοπνευμονική κόπωση
Λειτουργικός έλεγχος αναπνοής
Ανάλυση Αερίων Αρτηριακού Αίματος
Χρόνια αποφρακτική Πνευμονοπάθεια
Κυστική Ίνωση
Νεοπλάσματα Πνεύμονος ©
Παθήσεις διαμέσου Πνευμονικού ιστού
© κλιματική αλλαγή, άσθμα και αλλεργική ρινίτιδα
© κλιματική αλλαγή και πνευμονοπάθειες
10 αρχές για καθαρό αέρα
ενεργητικό και παθητικό κάπνισμα
https://d.pr/free/i/qubV1j
Ανάλυση Αερίων Αρτηριακού Αίματος, ΑΑΑΑ
Σύντομη προσέγγιση
Google Analytics: https://d.pr/free/i/OfcQtG

η παγκόσμια κίνηση του κοροναϊού
Κορονοϊός SARS-COV-2 και
η νόσος COVID-19.
2023: 32 πανελληνιο πνευμονολογικο συνέδριο -περιλήψεις ανακινώσεων
83: διαδερμική μέτρηση αερίων αίματος
Δενδριτικά κύτταρα
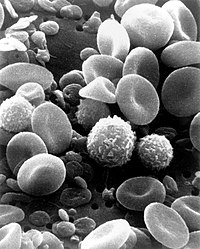
|&, &, &| Τα δενδριτικά κύτταρα σχηματίζουν το όριο μεταξύ του σύμφυτου και προσαρμοσμένου αμυντικού συστήματος. Η βασική τους λειτουργία έγκειται στην προώθηση αντιγόνων και την παρουσίασή τους στην επιφάνεια άλλων ανοσολογικώς δρώντων κυττάρων. Η ενεργοποίηση των ΔΚ από τα σχετιζόμενα με το παθογόνο αίτιο μοριακά δείγματα (=pathogen-associated molecular patterns, PAMP) είναι σημαντική για την πυροδότηση των πρωτοπαθών αποκρίσεων των λεμφοκυττάρων Τ. Τα ανώριμα δενδριτικά κύτταρα παραμένουν στους περιφερικούς ιστούς όπου εποπτεύον το περιβάλλον προκειμένου να εντοπίσουν σημεία λοιμώξεως. Με τον εντοπισμό παθογόνου, τα ΔΚ ενεργοποιούνται και μεταναστεύουν στους περιοχικούς λεμφαδένες όπου διαδραματίζουν κεντρικό ρόλο στην καθοδήγηση των Τ-λεμφοκυττάρων. Η διαδικασία της ενεργοποιήσεως των ΔΚ είναι πολύπλοκη και περιλαμβάνει διάφορα στάδια. Αμέσως μετά την ενημέρωση της παρουσίας PAMP στην περιφέρεια, τα ΔΚ υφίστανται παροδική αύξηση της ενδοκυτταρικής τους δραστηριότητας και προετοιμάζονται να προσλάβουν το ΡΑΜΡ και να μεταφέρουν την πληροφορία στα λεμφοκύτταρα στους περιοχικούς λεμφαδένες. Ακολούθως παρατηρείται ενίσχυση της εκφράσεως μορίων, όπως τα CD40 και CD86 και των επιπέδων του MHC κατηγορίας ΙΙ στην επιφάνεια του κυττάρου. Η ενεργοποίηση των ΔΚ επάγει, επίσης, την έκφραση χημοκινών, όπως η CCR7, που, με τη σειρά της διευκολύνει τη μετανάστευση των ΔΚ προς τους περιοχικούς λεμφαδένες, αμέσως μετά τη δειγματοληψία του μολυνθέντος κυτταρικού περιβάλλοντος. Η αρχική περιγραφή των δενδριτικών κυττάρων ανάγεται στον Paul Langerhans (κύτταρα Langerhans) στα τέλη του 19ου αιώνα, αλλά ο όρος ”δενδριτικά κύτταρα” οφείλεται στους Ralph M. Steinman και Zanvil A. Cohn. Για τη συμβολή του στην περιγραφή του κεντρικού ρόλου των δενδριτικών κυττάρων στο προσαρμοσμένο αμυντικό σύστημα στον Steinman απενεμήθη το βραβείο ”Albert Lasker”, το 2007 και το Βραβείο Nobel, το 2011[i].
βλέπε: φλεγμονή.
ο ρόλος των δενδριτικών κυττάρων στην καρκινογένενεση
Τα δενδριτικά κύτταρα διαδραματίζουν κρίσιμο ρόλο στην ενεργοποίηση υποπληθυσμών λεμφοκυττάρων (&), για τον έλεγχο ή τον περιορισμό των νεοπλασματικών ιστών. Με αναφορά στα νεοπλασματικά κύτταρα ή στα συνδεόμενα με τον όγκο αντιγόνα, τα δενδριτικά κύτταρα εκκολπώνονται τα αντιγόνα και μεταναστεύουν μέσω λεμφικών οδών στους περιοχικούς λεμφαδένες. Η πυκνότητα των άωρων μορφών δενδριτικών κυττάρων ( τα γνωστά ως κύτταρα Langerhans και δενδιτικά κύτταρα τιυ διάμεσου ιστού) και τα ώριμα δενδριτικά κύτταρα, παρόντα στο μικροπεριβάλλον του όγκουεμφανίζουν υψηλή προγνωστική αξία για το ειδικό εκ της πάθησης προσδόκιμο επιβίωσης ασθενών σε πρώιμα στάδια ΜΜΚΠ [2,3] καθώς, επίσης, η παρουσία τους στον χειρουργικά εξαιρέθεντα όγκο είναι καλόος προγνωστικός δείκτης [4]. Οι αντεπιδράσεις μεταξύ των δενδριτικών και καρκινικών κυττάρων απολήγουν στην απελευθέρωση αντινεοπλασματικών κυτοκινών [5,6], γεγονός που μπορεί να ερμηνευτεί ως έναρξη απαντήσεων από το προσαρμοσμένο αμυντικό σύστημα in situ [7, 8, 9]. Ο αριθμός και η λειτουργία των ώριμων δενδριτικών κυττάρων στο περιφερικό αίμα και τους λεμφαδένες ασθενών με ΚΠ, μειώνονται δραστικά, εν μέρει λόγω παθολογικής διαφοροποίησης των μυελοειδών κυττάρων [10, 11,12]. Τα καρκινικά κύτταρα και τα στρωματικά κύτταρα, όπως οι ινοβλάστες, και τα διηθούντα τους όγκους ανοσοκύταρα ή και τα απεκκρινόμενα προϊόντα τους, όπως ο VEGF, M-CSF,
IL-6, IL-10 και TGF-β απαντούν επίσης στα αλλείμματα των συστημικών και τοπικών δενδριτικών κυττάρων [13,14].
Τα προσβληθέντα δενδριτικά κύτταρα στερούνεται της ικανότητάς τους να φαγοκυτταρώνουν αντιγόνα, κια να ενεργοποιούν Τ-λεμφοκύτταρα, οδηγώντας σε έελλειμματική επαγωγή των αντινεοπλασματικών απαντήσεων. Τα δενδριτικά κύτταρα επί ΜΜΚΠ παράγουν υψηκές συγκεντρώσεις ογκοκατασταλτικής IL-10 και TGF-b [14]. Έχει δειχθεί ότι τα συνηανασταλτικό των Τ-κυττάρων μόριο B7-H3 και η λιγανδίνη-1 των υποδοχέων για τον προπγραμματισμένο θάνατο (PD-Li1) αναβαθμίζονται στα κατασκηνούντα στους όγκους δενδριτικά κύτταρα και τα μόρια αυτά εκπέμπουν μεγάλο αριθμό κατασταλτικών σημάτων, αναστέλλουν την παραγωγή κυτοκινών και Τ-κυττάρων.
ο ρόλος των δενδριτικών κυττάρων στην κυτταρική ανοσοθεραπεία. Τα δενδριτιικά κύτταρα είναι αντιγονο-παρουσιαστικά κύτταρα του αμυντικού συστήματος, που έχουν παλαιόθενα αναγνωριστεί ως ισχυοί ενεργοποιητές ανοσοαπαντήσεων. Λόγω της ικανότητάς τους να εκκολπώνονται αντιγονοπαρουσιαστικά κύτταρα και να ενεργοποιούν Τ-κύτταρα κατά ένα αντιγονο-ειδικό τρόπο, η χρήση δενδριτικών κλυττάρων ως ανσοθεραπευτικών παραγόντων έιναι ένας αξιοπρόσεκτος τομέας της σύγχρονης βιοθεραπευτικής. Τα δενδριτικά παράγονται ex vivo και , μετά τη διάστασή τουτς με σχετικά ογκογονίδια (respi-gam &), η επιστροφή τους στον ασθενή με την προοπτική αν αποκατασταθεί κατάλληλη αντιγονοπαρουσιαστική λειτουργία και ενεργοποίηση των Τ-κυττάρων. Η θεώρηση αυτή έχει διερευνηθεί στο ΜΜΚΠ και έχουν ληφθεί σημαντικά αποτελέσματα αναφορικά με την πρόκληση κατάλληλης ανοσο- απαντήσεως, σε συνθήκες ασφάλειας και ανοχής [17, 18, 19].
ανοσοπαθολογικές μεταβολές των δνδριτικών κυττάρων στη ΧΑΠ επί καπνιστών και μη καπνιστών.
[i] West JB. Ventilation / blood flow and gas exchange. Oxford: Blackwell, 1975.
2. Sautes-Fridman C et al (2011) Tumor microenvironment is multifaceted. Cancer Metastasis Rev 30(1):13–253.
3. Becker Y (1993) 3. Dendritic cell activity against primary tumors: an overview. In Vivo 7(3):187–191
4. Al-Shibli K et al (2009) The prognostic value of intraepithelial and stromal innate immune system cells in non-small cell lung carcinoma. Histopathology 55(3):301–312
5. Becker Y (1993) Dendritic cell activity against primary tumors: an overview. In Vivo 7(3):187–191
6. Mitra R, Singh S, Khar A (2003) Antitumour immune responses. Expert Rev Mol Med 5(3):1–19
7. Kusmartsev S, Gabrilovich DI (2006) Effect of tumor-derived cytokines and growth factors on differentiation and immune suppressive features of myeloid cells in cancer. Cancer
Metastasis Rev 25(3):323–331
8. Pinzon-Charry A, Maxwell T, Lopez JA (2005) Dendritic cell dysfunction in cancer: a mechanism for immunosuppression. Immunol Cell Biol 83(5):451–461
9. Shurin MR et al (2006) Intratumoral cytokines/chemokines/growth factors and tumor infi ltrating dendritic cells: friends or enemies? Cancer Metastasis Rev 25(3):333–356
10. Almand B et al (2000) Clinical signifi cance of defective dendritic cell differentiation in cancer. Clin Cancer Res 6(5):1755–1766
11. Bergeron A et al (2006) Characterisation of dendritic cell subsets in lung cancer microenvironments. Eur Respir J 28(6):1170–1177
12. Gabrilovich D (2004) Mechanisms and functional signifi cance of tumour-induced dendriticcell defects. Nat Rev Immunol 4(12):941–952
13. Steinman RM (1991) The dendritic cell system and its role in immunogenicity. Annu Rev Immunol 9:271–296
14. Dumitriu IE et al (2009) Human dendritic cells produce TGF-beta 1 under the infl uence of lung carcinoma cells and prime the differentiation of CD4 + CD25 + Foxp3+ regulatory T
cells. J Immunol 182(5):2795–2807
15. Mu CY et al (2011) High expression of PD-L1 in lung cancer may contribute to poor prognosis
and tumor cells immune escape through suppressing tumor infi ltrating dendritic cells maturation. Med Oncol 28(3):682–688
16. Schneider T et al (2011) Non-small cell lung cancer induces an immunosuppressive phenotype of dendritic cells in tumor microenvironment by upregulating B7-H3. J Thorac Oncol
6(7):1162–1168
17. Perroud MW Jr et al (2011) Mature autologous dendritic cell vaccines in advanced non-small cell lung cancer: a phase I pilot study. J Exp Clin Cancer Res 30:65
18. Wang K et al (2009) An autologous therapeutic dendritic cell vaccine transfected with total lung carcinoma RNA stimulates cytotoxic T lymphocyte responses against non-small cell lung cancer. Immunol Invest 38(7):665–680
19. Zhou Q et al (2008) A dendritic cell-based tumour vaccine for lung cancer: full-length XAGE-1b protein-pulsed dendritic cells induce specifi c cytotoxic T lymphocytes in vitro. Clin Exp Immunol 153(3):392–400
Μετρητής επισκέψεων

© Γεώ ργιος A. Mαθιουδάκης MD PhD
ργιος A. Mαθιουδάκης MD PhD
ΠNΕΥΜΟΝΟΛΟΓΙΚΟ ΚΕΝΤΡΟ ΑΘΗΝΩΝ
ATHENS BREATH CENTER
λ. Αλεξάνδρας 209, μετρό Αμπελοκήπων
 Αλέξανδρος Γ. Μαθιουδάκης MD, MRCP(UK) PhD
Αλέξανδρος Γ. Μαθιουδάκης MD, MRCP(UK) PhD
Λέκτορας Πνευμονολογίας, The University of Manchester & ΠNΕΥΜΟΝΟΛΟΓΙΚΟ ΚΕΝΤΡΟ ΑΘΗΝΩΝ
ISSN: 2241-4460
♦ΧΡΟΝΙΑ ΑΠΟΦΡΑΚΤΙΚΗ ΠΝΕΥΜΟΝΟΠΑΘΕΙΑ, ΧΑΠ
♦ΑΣΘΜΑ
♦ΠΝΕΥΜΟΝΙΚΕΣ ΕΚΔΗΛΩΣΕΙΣ ΕΞΩΠΝΕΥΜΟΝΙΚΩΝ ΠΑΘΗΣΕΩΝ
♦Πνευμονική αποκατάσταση